



5. Онтология и методология теории Н.А. Бернштейна
Основное содержание онтологической картины концепции Н.А. Бернштейна зафиксировано в самом ее названии — «физиология активности». Если действующие лица, на первый взгляд, остались здесь прежними (Н.А. Бернштейн так же, как и И.П. Павлов, рассматривает схему «организм-среда»), то содержание ролей радикально изменилось: понятию организма и его основному отношению к среде даются совершенно новые интерпретации.
В то время как традиционная физиология, занимаясь поведением, ограничивалась, по существу, рассмотрением отдельных приспособительных актов, в теории Н.А. Бернштейна горизонт физиологического умозрения расширяется до анализа жизни особи. Для этого потребовалось в первую очередь пересмотреть сложившееся понятие организма, трактовавшее его как реактивно-уравновешивающуюся систему.
В концепции Н.А. Бернштейна организм рассматривается как организация, характеризующаяся двумя главными свойствами. Во-первых, это организация, сохраняющая свою системную тождественность сама с собой, несмотря на непрерывный поток как энергии, так и вещества, субстрата, проходящих через нее. Несмотря на то, что ни один индивидуальный атом в организме не задерживается в составе его клеток, организм остается сегодня тем же, чем был вчера, и его жизнедеятельность обусловливается всей его предшествующей жизнью.
Во-вторых, — развивает биологическую диалектику Н.А. Бернштейн, — организм на всех ступенях и этапах своего существования непрерывно и направленно изменяется. Эта направленность онтогенетической эволюции неоспоримо доказывается хотя бы тем, что тысяча представителей одного животного или растительного вида развивается в особей, одинаковых по своим основным или определяющим признакам, несмотря на иногда весьма резкую неодинаковость внешних условий жизни у разных индивидов. Что касается эмбриогенеза, то, начиная уже со стадии оплодотворенного яйца, организм обладает закодированной моделью будущего своего развития, оформления и закодированной же программой последовательных ступеней этого развития.
Самое же важное, по мнению Бернштейна, состоит даже не в этой «запрограммированности», а в том динамическом начале (в конце концов, вероятно тоже как-то закодированном и обладающим своим вещественным субстратом-носителем в клетке), которое создает у особи активное антиэнтропийное, преодолевающее стремление к реализации этой кодированной модели (Бернштейн, 1963,с. 313).
Какие следствия влечет за собой это изменение понятия организма для онтологической картины концепции? Ясно, что отмеченная выше тождественность результатов морфогенетического развития на фоне изменчивых условий говорит о том, что организм активно преодолевает возможные и неизбежные внешние препятствия на пути программы своего морфогенеза. Экспериментальные факты повреждений и частичных ампутаций (например, конечностей) в эмбриогенезе, ампутаций, не мешающих этим органам развиваться в полноценную конечность; факты анатомических, а еще более функциональных регенераций; клинический материал — все эти данные говорят о том, что организм активно борется за свое выживание, развитие и размножение. Процесс жизни — это не уравновешивание с окружающей средой, как считал И.П. Павлов. Такое уравновешивание обрекло бы каждую особь на полную зависимость от среды и ее изменений, в результате чего о программном морфогенезе с удержанием стойких признаков вида нельзя было бы и думать. Процесс жизни — это преодоление среды, направленное при этом не на сохранение статуса или гомеостаза, а на движение в направлении родовой программы развития и самообеспечения (Бернштейн, 1963, с. 314).
Такому общему взгляду на жизнь особи, на отношение «организм—среда», важнейшее в онтологии биологических дисциплин, полностью соответствует произошедшая в концепции Н.А. Бернштейна смена представлений об отдельном акте поведения, рассмотренная выше. Он существенно определяется средой, но никогда полностью не детерминирован ею. Активность — эта важнейшая черта живых систем (Бернштейн, 1966, с. 188) — на уровне отдельных поведенческих актов проявляется в их целеустремленности. Если понятие цели в концепции И. П. Павлова, как мы видели, в лучшем случае допускалось лишь в исследовательскую «кухню» как эвристический познавательный прием, то в физиологии активности Н.А. Бернштейна это понятие онтологизируется. Без него факты сложного поведения животного, факты координации и управления собственными движениями попросту необъяснимы.
Обнаружение и исследование «сложности движения» в своем методологическом звучании явилось открытием предметного характера движения. Тривиальный, но от этого не становящийся несущественным факт активного взаимодействия живого существа с вещами внешнего мира совершенно не учитывался павловской физиологией. Она видела, с одной стороны, движение (сводимое к реакции), с другой — предмет (сводимый чаще всего к раздражителю, условному или безусловному), но не видела живого предметного движения, то есть движения, во-первых, удовлетворяющего некоторую жизненно важную потребность организма, а значит, в определенных своих параметрах жестко заданного этой потребностью, и, во-вторых, развертывающегося в предметном мире и вынужденного поэтому, чтобы быть успешным, соответствовать по своему составу объективным свойствам этого мира.
Сложное строение тела высшего животного, с одной стороны, и подвижное многообразие объективных обстоятельств, с другой, делают всякую поведенческую ситуацию практически уникальной, так что даже самый богатый репертуар врожденных реакций в принципе не мог бы обеспечить каждую из ситуаций адекватным движением. Движение должно быть всякий раз заново построено.
Нужно было коренным образом изменить представление об организме и способе его жизни, как это сделал Н.А. Бернштейн, чтобы поставить в конкретно-научной плоскости проблему построения движений. Ее решение с логической необходимостью потребовало перестройки концептуального аппарата физиологической науки в той его части, которая была предназначена для изучения поведения. Сюда относится переход от схемы рефлекторной дуги к схеме рефлекторного кольца, развитие представлений о коррекции движений с помощью проприорецепторики (в широком смысле), что означало открытие важнейшей контрольно-корректировочной функции афферентации наряду с открытой И.П. Павловым сигнально-пусковой ее функцией. Наконец, в аппарат поведенческих дисциплин был введен ранее считавшийся крамольным комплекс понятий: цель, задача, «модель потребного будущего». Эти понятия, фиксируя важнейший механизм реализации сложного поведения, явились в то же время конкретным воплощением общей идеи активности — активного, деятельного, преодолевающего отношения организма к среде, — пришедшей на смену представлениям об этом отношении как об уравновешивании.
Столь значительный прогресс в теоретических представлениях был возможен лишь при овладении новыми, более мощными методологическими средствами. В этой методологической плоскости концепцией Н.А. Бернштейна был сделан шаг не меньший, чем в плоскости конкретно-научной. В науках о поведении это был шаг от методологии натуралистической к методологии деятельностно-ориентированной.
Центральная для методологии науки проблема детерминизма была в физиологии активности решена парадоксальным для классического физиологического натурализма образом: не столько законы функционирования мозговой ткани определяют поведение животного, то есть его реальную жизнь, сколько сама эта жизнь, деятельное решение жизненных задач определяет функционирование мозга, а в эволюционных масштабах и его строение.
Можно сказать, что Н.А. Бернштейн открыл для конкретно-научного физиологического исследования поведенческую реальность, сумев описать его единицу — «живое движение». Входящий в состав живого движения механизм «сенсорных коррекций» делает его движением «умным», «зрячим», поэтически выражаясь — «исполненным очей», а психологически — осмысленным, изнутри просветленным отражением предметной реальности. Но само это отражение, этот «ум» живого движения вовсе не самостоятельная, отдельная от собственно движения инстанция, а «функциональный орган» (см. Зинченко, 1997; Зинченко, Гордеева, 1982), вне и помимо самого движения не работающий. Все это известные и житейскому самонаблюдению вещи: так нога, нащупывая в темноте тропинку, одновременно и исполняет очередной шаг, и изучает предметную реальность, и гибко перестраивает свое движение в соответствии с этой реальностью. Впрочем, то, что для житейского сознания просто, то нередко чрезвычайно сложно для точной научной фиксации.
Это, так совершенно устроенное у высших животных, внешнее поведение, в павловской теории, несмотря на ее претензии объяснить фундаментальные законы поведения, по существу изгонялось из физиологической науки. Реальное поведение (как действие, так и восприятие) низводилось в павловской системе до роли источника информации для мозга животного и индикатора гипотетических мозговых процессов (индукции, иррадиации, замыкания и т.п., что Б.Ф. Скиннер ядовито, но точно назвал «концептуальной нервной системой») для исследователя.
Ничего интересного от исследования строения самого поведения не ждали, весь интерес был направлен вглубь, за черепную коробку, содержащую, как думалось, в себе все тайны и законы поведения животного и человека.
Гносеологическая, лабораторная редукция поведения до роли источника информации невольно онтологизировалась, так что основной жизненной задачей животного считались не действия, а ориентирование в среде, получение своевременной информации о появившихся раздражителях. (Да и то сказать, если весь репертуар врожденных реакций всегда в распоряжении животного, главное — своевременная информация, точная ориентировка, а уж за правильным приказом и его точным исполнением дело не станет.)
Эта «информационно-ориентировочная» парадигма, заданная павловской методологией, продолжала реализовываться в трудах его учеников и последователей, в первую очередь в теории П.К. Анохина, несмотря на, казалось бы, радикальные преобразования, внесенные этой теорией в павловские представления о поведении, — понятие акцептора действия, принцип обратной связи и пр. И все же, несмотря на бесспорную продуктивность и на новейшую для того времени кибернетически-информационную терминологию, эти теории лишь закрепили главный методологический порок павловского учения — неспособность научно увидеть самостоятельную, полновесную реальность живых, предметных двигательных актов, несводимых ни к каким условным сигналам, ни к какой информационной представленности в мозгу.
Кибернетического толка теории поведения при неоправданной экспансии информационных абстракций склонны к замещению реальности информацией о реальности, как в физиологическом мышлении И.П. Павлова реальное событие — раздражитель — легко замещалось нервным возбуждением афферентной клетки, а реальное движение — возбуждением клетки эфферентной (вспомним павловский тезис: «Последняя инстанция движения в клетках передних рогов спинного мозга»). Будь этот биологический иллюзионизм справедлив, для боксера было бы безразлично, получает ли он информацию об ударе или удар, для экспериментального животного — получает ли оно информацию о пище или саму пищу. Корыстолюбивый предтеча такой информационной теории взаимодействия живого существа с миром, хозяин стамбульской харчевни требовал от бедняка расплатиться за запах шашлыка, но Ходжа Насреддин указал ему на методологическую непоследовательность, предложив получить плату не монетами, а звоном монет.
Но, возвращаясь к физиологии активности, можно спросить: разве сам НА. Бернштейн, особенно в последних своих работах, не использовал в качестве ключевой идею обратной связи и не подошел к информационно-кибернетической методологии? Использовал и подошел, но стоит полностью согласиться с квалификацией А.Н. Леонтьевым этой переориентации взглядов Н.А. Бернштейна как «известного отступления от ранних работ, в которых развивался принцип активности…» (Леонтьев, 1972, с. 79). Дело в том, что представление о кольцевой регуляции «живого движения» с помощью сенсорных коррекций, осуществляемых на разного уровня «сенсорных полях», хотя и содержит в себе с формально-логической точки зрения одну из центральных идей кибернетики — принцип обратной связи, но для исследования поведения намного продуктивнее и богаче этой идеи и сведена к ней быть не может.
Понятие обратной связи в физиологии поведения (реализованное, в частности, в концепции акцептора действия П.К. Анохина) оперирует поведенческими событиями как готовыми, атомарными сущностями. Движение при этом попадает в поле зрения теории только до его начала (в виде цели) и после его завершения (в виде результата). Представление же о сенсорных коррекциях и сенсорных полях разных уровней ухватывает движения животного в его живом протекании, в его динамическом построении, которое вовсе не является суммой атомарных проб исполнения данного движения. Не случайно главный труд Н.А. Бернштейна так и назван «О построении движений». Поэтому свести методологию «физиологии активности» к идее обратной связи — значит пройти мимо самого интересного, творческого, глубокого и перспективного в ней.
Итак, на уровне уже философской, а не собственно методологической рефлексии теория условных рефлексов репрезентирует философию натуралистического иллюзионизма.
Физиология активности, мыслящая поведенческий акт как живой орган встречи с реальностью, функционирование которого изнутри просветлено перцептивным отражением этой реальности, а сама эта перцепция опосредована реальным предметным движением, — это теория, которая воплощает совсем другую философию — философию энергийного реализма.
2.3. Павлов и Скиннер: сравнительный методологический анализ теорий
Название главы можно было бы стилизовать под логическую формулу «(Павлов & Скиннер) V Бернштейн?», пытаясь так выразить, во-первых, утверждение, что истинно одно из двух — либо теория Н.А. Бернштейна, либо концепции И.П. Павлова и Б.Ф. Скиннера вместе взятые, а во-вторых, вопрос — что же все-таки истинно? Однако от подобного заголовка пришлось отказаться потому, что о теории И.П. Павлова речь пойдет только в самом конце главы, а фамилия Н.А. Бернштейна и его конкретные теоретические идеи почти вовсе не будут фигурировать на последующих страницах; но, главное, потому, что для автора эта дизъюнкция уже потеряла свой вопросительный знак, она решена в пользу Н.А. Бернштейна. Авторская точка зрения совмещена в рамках данной работы с той методологической и теоретической позицией, которую символизирует имя Н.А. Бернштейна, и анализ оперантного бихевиоризма Б.Ф. Скиннера ведется с опорой именно на это основание, само по себе остающееся вне обсуждения, как бы вынесенным за скобку. Что касается знака конъюнкции между Павловым и Скиннером, то он использован здесь скорее в психологическом, чем в логическом смысле и выражает сложившееся у автора убеждение, что концепции этих ученых вырастают из одного и того же методологического корня, имеют один и тот же философский и методологический «генотип», черты которого, несмотря на все «фенотипические» различия, явно проступят, если поставить эти две теории рядом.
«Поставить теории рядом» — значит придать им сопоставимую форму, чтобы можно было сравнивать не по их самопредъявлению, не по поверхностным признакам, а по внутренней сути. Для этого нам понадобится методологически препарировать концепцию радикального бихевиоризма так же, как это было сделано в предыдущей главе по отношению к теории условных рефлексов и с помощью того же, уже знакомого читателю методологического аппарата (онтология — основной идеальный объект — предмет — объект — метод).
Радикальный бихевиоризм Б.Ф. Сканера
1. Онтология
Онтологией радикального бихевиоризма является та же, что и у И.П. Павлова, схема «организм—среда». Однако понятие организма здесь пересмотрено, точнее — принципиально и сознательно недосмотрено: исходя из своей позитивистской установки, Б.Ф. Скиннер отказывается от учета всех внутренних, во внешнем поведении непосредственно не наблюдаемых процессов — физиологические они или психологические — неважно, а вернее, неизвестно. Организм есть «черный ящик», некая непрозрачная емкость, в принципе не проницаемая, а главное — неинтересная для бихевиористского взгляда. Организм — это место, в котором нарушается непрерывность наблюдаемых в онтологии «организм—среда» процессов, это как бы дыра, брешь в среде, где бесследно исчезают и откуда неожиданно появляются наблюдаемые процессы. Между теми и другими, интерпретируемыми соответственно как стимулы и реакции, естественно, устанавливаются не причинные отношения, предполагающие наличие непрерывности, а отношения корреляционные. Скиннер «игнорирует возможность промежуточных физиологических звеньев… — пишет известный историк психологии Е. Боринг. — Такие функциональные отношения, как R = f(S), устанавливаются наблюдением за соизменениями S и R и лишены физической непрерывности между терминами, которую предпочитают большинство ученых. Скиннеровские функции — просто корреляции дискретных переменных, и друзья Скиннера порой шутят, что он имеет дело с пустым организмом» (Boring, 1950, р. 650).
В шутке этой, впрочем, шуточного немного, она является вполне точной констатацией ядерной методологемы радикального бихевиоризма, которую можно назвать абстракцией «пустого организма». Эта абстракция чрезвычайно последовательно проводится Скиннером на всех уровнях исследовательской работы, начиная от решения методологического вопроса об отношениях между физиологией и психологией и кончая способом описания и фиксации конкретных экспериментальных условий. Нелепо было бы, конечно, понимать эту абстракцию буквально. Разумеется, Скиннер знает, «что организм не пуст и поэтому не может быть адекватно изучен просто как "черный ящик", но, — настаивает он, — мы обязаны внимательно относиться к разнице между тем, что нам действительно известно о находящемся внутри, и тем, что просто логически выводится» (Skinner, 1974, р. 213). Абстракция «пустого организма» — лишь квазионтологическое выражение пафоса гносеологической борьбы Скиннера против метафизических демонов и гомункулусов, в какие бы материалистические одежды они ни рядились в теориях современных физиологов и психологов, приписывающих своим концептам реальную силу и способность упорядочивать поведение и управлять им. Другими словами, абстракция «пустого организма» — выражение неудовлетворенности «менталистской» психологией (считающей возможным ссылаться при объяснении поведения на такие ненаблюдаемые вещи, как желание, потребность, намерение и т.п.) и «концептуальной» физиологией, апеллирующей к столь же мистическим сущностям. Дело в том, поясняет Скиннер, что с античности поведение объясняется смесью анатомических, физиологических и менталистских фактов. «Его сердце разбито в любовной драме»; «Он ошибся, потому что его нервы были натянуты» и т.д. и т.п. Это обстоятельство сказалось не только на психологии, но и на физиологии. «Были выделены различные части нервной системы, но что происходит в каждой из них — это только логически выводилось. Отчасти такое положение сохранилось и в XX веке. Синапс, проанализированный сэром Чарльзом Шерингтоном, был частью концептуальной (читай «воображаемой». — Ф.В.) нервной системы; то же относится и к "деятельности коры больших полушарий", исследованной Павловым» (Skinner, 1974, р. 213). Чем пользоваться воображаемой нервной системой «для объяснения поведения, из которого она сама логически выводится», — считает Скиннер (там же), — лучше уж вообще отказаться пока от всякого физиологического объяснения.
Хотя в отношении будущей физиологии Скиннер настроен чуть более оптимистично, но и ей отводится весьма скромная роль заполнения бреши между стимулами и реакциями, оставленной ей бихевиоризмом, знаниями о химических и электрических процессах, которые происходят в организме, когда он действует (Skinner, 1931, 1974). Впрочем, и тогда «то, что откроют физиологи, не сможет сделать недействительными законы, установленные наукой о поведении» (Skinner, 1974, р. 215).
При всей справедливости критики Б.Ф. Скиннером физиологических фантазий нельзя не заметить, что он принципиально вычеркивает физиологию из списка поведенческих дисциплин, хотя из вежливости и говорит, что в будущем, когда она наберется ума, ей, может быть, и доверят закрасить белые пятна на карте научного объяснения поведения, но уж, конечно, никогда не доверят самой рисовать контуры поведенческого ландшафта. В самом деле, если, по мнению Скиннера, никакое самое совершенное физиологическое знание не сможет поколебать уже установленных законов поведения, значит, исторически сложившаяся граница между физиологией и бихевиоральной психологией возводится в ранг принципа, а следовательно, в онтологической плоскости строится непреодолимая стена между внешним и внутренним (физиологическим), между поведением организма и организмом. Поэтому психолог-бихевиорист, хотя и может помечтать, что физиологи когда-нибудь расскажут ему, «что происходит, когда, например, ребенок учится пить из чашки», или более того, «как, следя за изменениями в нервной системе, добиться, чтобы он научился это делать» (Skinner, 1974, р. 214), но в качестве профессионала он должен забыть, что за кожей что-то есть, что там происходят процессы, существенно сказывающиеся на поведении, или — упаси Бог — что есть нечто внутреннее, которое и в поведении не наблюдается, и в организме самым совершенным прибором не сыщешь и которое тоже вносит свой вклад в организацию и осуществление поведения. Хочешь оставаться ученым — считай, что «организм пуст», ищи детерминанты поведения в видимой внешней среде, отказывайся от метафизической привычки ссылаться на какое-нибудь «переключение возбуждения» или «намерение», которых никто никогда не видел и которых, следовательно, не существует как научных фактов. Вполне очевидно, что на философском уровне радикальный бихевиоризм является радикальным позитивизмом.
Приведем конкретный пример реализации позитивистских заповедей в познавательной практике радикального бихевиоризма. Когда в экспериментах Скиннера используется пищевое подкрепление, вес животного доводится, например, до 75% от обычного веса. Дело здесь не в том, что Скиннер хочет дать некоторое объективно фиксированное и потому операционально воспроизводимое выражение потребности, в данном случае пищевой. Он в принципе отказывается от понятия потребности (как бы его ни трактовали — физиологически или психологически) как некоего внутреннего состояния организма, являющегося причиной поведения. Здесь нет предположения, что у всех участвующих в эксперименте животных (или у одной особи в разное время) при доведении их веса до 75% от нормы пищевая потребность будет одинакова. Потребность, по Скиннеру, не есть то, что означается или выражается в этом весе, она есть 75% веса, и ни о какой другой таинственной потребности, приписываемой внутреннему миру организма, говорить нельзя, если мы хотим говорить научно (разумеется, в позитивистском смысле этого слова).
Скиннер утверждает: раз вы не можете объективно наблюдать потребность саму по себе, нельзя объяснять ею поведение, ибо само понятие потребности вы сначала выводите из поведения, а затем им же это поведение объясняете. Это физиологическая или психологическая мистификация, от которой бихевиорист должен отказаться. Когда вес животного перед экспериментом доводится до 75%, то это — некоторый твердый факт, имеющий определенную фиксацию. Если при этом утверждать, что животное испытывает голод и потому действует, то это ровным счетом ничего не прибавляет к нашему знанию и, главное, никак научно, операционально не может быть учтено. Так что в экспериментальном мышлении Б.Ф. Скиннера не имеется в виду, что есть самостоятельная сущность — «потребность в пище», которая в данном эксперименте количественно выражается «семидесятипятипроцентным весом животного». Есть просто эти 75% веса, и, если угодно, можете называть это потребностью, но не вкладывайте в это слово ваших привычных ассоциаций, за ним ничего не должно стоять, кроме указанных процентов и соответствующих процедур взвешивания.
Так «организм» в онтологии радикального бихевиоризма последовательно очищается от всех внутренних содержаний. Что касается понятия среды в этой онтологии, то его удобнее обсудить несколько позже.
2. Основной идеальный объект радикального бихевиоризма — оперантный рефлекс
Б.Ф. Скиннер так же, как и И.П. Павлов, полагает, что жизнь организма осуществляется за счет безусловных и условных рефлексов, но в отличие от русского исследователя он выделяет два типа обусловливания — классическое, или респондентное, изучавшееся в павловской школе, и оперантное обусловливание. Рассмотрим основные различия между этими типами. Б.Ф. Скиннер так схематизирует их (Skinner, 1935) :
|
|
|
Схема 1. Различия оперантного и респондентного обусловливания (приводится с несущественными изменениями по изданию — Skinner, 1959, р. 367) |
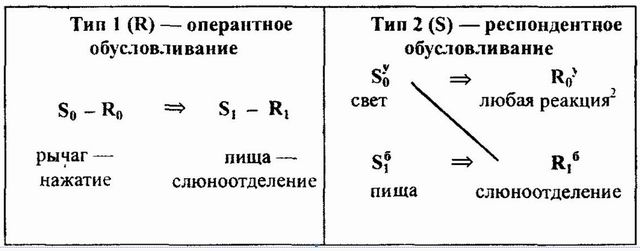
Центральное различие между этими типами приходится на результаты обусловливания. В первом случае происходит изменение силы рефлекса (S0 — R0), которая измеряется частотой или вероятностью появления ответа R0 при наличии стимула S0; во втором типе устанавливается условная связь S0y — R1б, то есть возникает новый рефлекс. Иначе говоря, оперантное обусловливание не порождает новых рефлексов, оно только увеличивает или уменьшает (в зависимости от того, положительное подкрепление или отрицательное) их силу, при респондентном же обусловливании возникает новый рефлекс, в котором ранее безразличный раздражитель S1б занимает место безусловного S1б и обретает способность вызывать соответствующую безусловную реакцию R1б.
Чрезвычайно важны для характеристики обоих типов временные условия подкрепления. Чтобы произошло оперантное обусловливание, подкрепление S1 должно появиться после того, как произошла реакция R0. Во втором типе для обусловливания существенным является временное отношение подкрепления S1б не с реакцией R0у, а с безразличным раздражителем S0у. Чтобы стать условным, стимул S0у должен предшествовать или появляться одновременно с безусловным раздражителем (подкреплением) S1б.
В статье «Два типа условных рефлексов и псевдотип» (Skinner, 1935) Скиннер проводит еще целый ряд различий между R- и S-обусловливанием, но для целей нашего методологического анализа достаточно и только что описанных.
Коснемся вкратце вопроса о возможности сведения двух типов обусловливания друг к другу. К чести Скиннера надо сказать, что он избегает соблазна сводить классическое обусловливание к оперантному (Skinner, 1935). Правда, он утверждает, что чистый условный рефлекс эмпирически получить невозможно ввиду того, что во время подкрепления на животное действует не один только предназначенный стать условным раздражитель (например, свет), но вся совокупная стимульная ситуация. Поэтому для образования условного рефлекса на данный стимул требуется введение дополнительных условий, в самой схеме классического обусловливания не учитываемых, которые обеспечивали бы выделение S0у из стимульного поля, что экспериментально достигается его неожиданным появлением (Skinner, 1935). С точки зрения павловской теории условных рефлексов это просто недоразумение, основанное на игнорировании роли ориентировочной реакции: сначала Скиннер принимает, что для образования «чистого» условного рефлекса респондентного типа не существенна первичная реакция R0у на безразличный стимул, то есть ориентировочная реакция (там же), а затем, исходя из того, что для установления условной связи S0у — R1б требуется некоторая активность организма по выделению S0у из стимульного поля, он утверждает, что «чистого» условного рефлекса не существует. Скиннер почему-то отказывается допустить возможность, что та самая реакция R0у, которой он пренебрег в схеме образования классического условного рефлекса, как раз и берет на себя функцию выделения стимула. Почему же? Ведь экспериментально, в чисто бихевиористской манере не так уж трудно зафиксировать, например, снижение порогов восприятия объекта, который стал стимулом ориентировочной реакции. Но тем не менее придание должного функционального значения ориентировочной реакции привело бы Скиннера к недопустимым для радикального бихевиоризма выводам о наличии перцептивного взаимодействия животного со средой, изменяющего эту среду не материально, а идеально — приданием тем или иным объектам особого статуса в жизненном пространстве животного.
Что касается обратной возможности — сведения оперантного обусловливания к респондентному, то павловская школа вообще отказывается признать, что Скиннером (точнее, Миллером и Конорским) был открыт новый тип обусловливания, утверждая, что он без остатка сводим к закономерностям образования классического условного рефлекса. Ход рассуждения базируется при этом на отождествлении оперантной реакции Rg с безразличным стимулом Sg": собственное движение животного может явиться для него точно таким же стимулом, как и любое другое событие, и, следовательно, может быть рассмотрено как раздражитель. А раз этот раздражитель предшествует появлению подкрепления, то со временем он становится условным, то есть начинает вызывать соответствующую подкреплению безусловную реакцию. Ни логически, ни фактически это рассуждение не противоречиво: если нажатие на рычаг точно так же, как и появляющийся независимо от животного звонок, сопровождается кормлением, и то, и другое в равной мере могут вызвать слюноотделение. Но тем самым просто усматривается функционирование классического обусловливания в ходе и во время оперантного обусловливания. А это вовсе не означает сведение последнего к респондентному, поскольку основной факт оперантной теории — увеличение вероятности оперантной реакции R при положительном подкреплении здесь не объясняется. Этот факт в стандартной схеме павловского эксперимента был бы равнозначен — смешно сказать — увеличению вероятности зажигания лампочки по той причине, что свет от нее стал для животного условным раздражителем.
По-видимому, разумнее всего признать (как это и делает Скиннер — Skinner, 1935), что работают всегда оба механизма, и усмотрение функционирования одного во время действия другого не устраняет ни тот, ни другой.
Рассмотрим теперь само понятие оперантного рефлекса безотносительно к его сопоставлению с респондентным.
Как изменяется оперантный рефлекс в результате подкрепления?
Существенной особенностью оперантного обусловливания является отсутствие подкрепления до тех пор, пока не произойдет оперантная реакция (Skinner, 1935), то есть последняя — это реакция на биологически нейтральный раздражитель и может быть названа поэтому «безразличной» реакцией по аналогии с понятием безразличного раздражителя у Павлова. Другими словами, до первого подкрепления оперантная реакция осуществляется, не «имея в виду» своих биотических последствий, вне какой-либо данности животному жизненно важного объекта и, соответственно, вне данности связи между его движениями и каким-то их будущим значимым результатом. Однако и после того, как произошло обусловливание, ошибочно утверждать, что животное «прозрело» относительно последствий подобного движения в сходных условиях и что оно будет осуществлять его в следующий раз потому, что «предвосхищает» или, того хуже, — «надеется» на получение такого же результата. Все это, — говорит Скиннер, — не более чем менталистские домыслы. Обученная крыса нажимает на рычаг в скиннеровском ящике вовсе не потому, что «ожидает», будто это приведет к появлению пищи… Просто в результате подкрепления увеличивалась вероятность такой реакции при наличии подобного стимула. Человек в таком изменившемся состоянии может переживать, что он нечто «предвосхищает», «ожидает», но это эпифеномен, необходимости в таком переживании нет, психическая деятельность не существенна для оперантного поведения (Skinner, 1974).
В результате оперантного обусловливания нового рефлекса не появляется, а в старом не происходит никаких содержательных преобразований, никаких качественных перестроек, изменяется лишь его сила, или вероятность его появления (Skinner, 1935).
Какова связь между стимулом и ответом в рефлексе?
Стимулом Скиннер называет некоторое воздействие среды, «вызывающее энергетическое изменение на периферии» (Skinner, 1931). Реакция (ответ) тоже есть некоторое внешнее событие, движение, доступное наблюдению. Естественно, что стимул не может рассматриваться как причина реакции, ибо наблюдение непрерывной причинной цепи в организме прерывается, и Скиннер не считает необходимым или даже полезным для анализа поведения знать о том, что происходит внутри, за кожей. Ссылаясь на Э. Маха, он заменяет понятие причинения стимулом реакции понятием функционального отношения между ними (там же) и ставит задачу выявления корреляций: связь между ними в рамках радикального бихевиоризма не онтологизируется, она, как уже говорилось, отдается на откуп физиологии, которая, по мнению Скиннера, и должна заполнять брешь, оставляемую ей бихевиоризмом (Skinner, 1974), «стремясь к описанию рефлексов в терминах физико-химических событий» (Skinner, 1931, р. 336).
Итак, стимул рассматривается Скиннером не как причина, а как фактор (если дать понятию фактора определение детерминанты, про которую известно, что она влияет на наблюдаемые события, но содержательный характер действия которой не является ни известным, ни искомым). «…Стимулы, — утверждает Скиннер, — не вызывают оперантных ответов, они просто изменяют вероятность того, что эти ответы произойдут» (Skinner, 1974, р. 223). Они обладают этой способностью не сами по себе, а в силу того, что присутствуют во время действия подкрепляющих обстоятельств.
Какова должна быть связь между оперантной реакцией и подкреплением для того, чтобы произошло обусловливание?
Исходя из здравого смысла можно было бы сказать, что подкрепление данного акта поведения происходит потому, что он производит некоторые преобразования среды или положения тела животного в среде так, что этим обеспечивается удовлетворение той или иной потребности. Однако такое рассуждение в корне противоречит духу скиннеровской теории. Во-первых, «ошибочно говорить, что пища оказывает подкрепляющее действие потому, что мы чувствуем голод, или потому, что мы чувствуем потребность в пище» (Skinner, 1974, р. 50). Чувство голода есть лишь ощущение некоторого условия, участвующего в процессе подкрепления, причем это условие действует вне зависимости от того, ощущается оно или нет. Во-вторых, — и это главное — связь между реакцией и возникающим после нее безусловным стимулом, подкрепляющим ее, является не предметной, содержательной связью, а отношением временного следования. Они могут быть, конечно, связаны и содержательно-предметно, когда оперантная реакция является предметной причиной возникновения в стимульном поле животного подкрепления, но таковой эта связь будет лишь по совпадению, а не по существу. С точки зрения скиннероаской схемы оперантного обусловливания реальный характер связи между реакцией и подкреплением (и формы чувственной данности животному этой связи) несущественен, то есть теоретически не различим (хотя эмпирически он, конечно, вполне может быть зафиксирован). Реакция в этой схеме будет подкреплена потому, что безусловный стимул последовал за ней, а не потому, что она его вызвала.
Оперантная реакция (ответ)
Центральным звеном, сердцем радикального бихевиоризма является представление об оперантной реакции. Если в результате подкрепления оперантный рефлекс S—R не появляется, не исчезает и никак не перестраивается, а лишь увеличивается или уменьшается вероятность его появления, значит и часть его, реакция R, в прижизненном опыте животного не испытывает никаких преобразований. Оперантная реакция есть, таким образом, врожденное, твердое, не изменяющееся в онтогенезе двигательное образование. В этом своем готовом виде она время от времени «выбрасывается» организмом в среду и затем, как пружина, вновь возвращается в исходное положение. Она, так сказать, предлагает процессам приспособления принять себя такой, как она есть, и только назначить ей ту или иную частоту своего проявления при наличии определенной стимульной ситуации. Подобно герою авантюрного романа (см. Бахтин, 1975), она возвращается после столкновения с предметным миром в том же виде и состоянии, в котором ушла, ни на йоту не изменившись, а только испытав и удостоверив в этом столкновении свой неизменный состав. Реакция не деформируется и не преображается, в ее фактуре не остается никаких осадков, примесей и следов от ее выхода в свет. Это представление мы будем условно называть абстракцией «чистого движения»( С равным правом ее можно было бы назвать и абстракцией «готового», «твердого», «атомарного» или «генотипического» движения. Эта абстракция является одним из вариантов общей и коренной для всего бихевиоризма и рефлексологии абстракции «непредметного движения». Но о6 этом — речь впереди.).
Если пренебречь некоторыми тонкостями, можно сказать, что наблюдать «чистые» движения мы могли бы только имея оптический прибор, вычитающий все влияния, которые оказываются на эти движения со стороны внешних предметов. В реальном же эмпирическом наблюдении мы имеем дело всегда с «фенотипом» данной реакции, который есть равнодействующая его врожденного состава и сил внешней среды.
Здесь, в этом пункте своей концепции, Скиннер сталкивается с самой, вероятно, сложной теоретической и методической проблемой — проблемой идентификации данного оперантного ответа. Она должна рассматриваться в двух аспектах — во-первых, как проблема отождествления нескольких в разное время происходящих реакций, во-вторых, как проблема временных границ, начала и конца данной оперантной реакции.
На идентификации разновременных реакций основывается весь массив экспериментальных исследований радикального бихевиоризма. В самом деле, если экспериментатор должен оценить изменение вероятности появления реакции, он должен быть уверен, что наблюдаемая им сегодня реакция животного есть та же самая реакция, которую животное осуществляло вчера. Предположим, в эксперименте исследуется оперантная реакция нажатия на рычаг, причем подкрепляются только нажатия с определенной силой F. Такая реакция и будет искомым оперантным ответом Rf. Если самописец, фиксирующий силу нажатия, достигает отметки F, значит произошла данная реакция Rf. Но все дело в том, что животному доступно практически бесконечное число движений R1, R2 R3, … Rn, с помощью которых можно произвести одинаковое нажатие экспериментального рычажка. Схематически в векторном виде это можно изобразить так:
|
Страница сформирована за 1.31 сек
SQL запросов: 191 |



